

























The Mystery of the Brain (Consciousness Series: Part 2)
- Oct 6, 2019
- 9 min read
Last blog post, we dove a bit into the complexity of consciousness and an introduction to various topics of consciousness research. This week we will delve a bit more into the technical side of consciousness research. How can one measure and assess consciousness, specifically in people with disorders of consciousness? How does neuroimaging play a role in the field of consciousness research? And, most importantly, how can we even begin to conceptualize consciousness from a neurological standpoint?
Neural Bases of Consciousness:
Let’s begin our discussion by defining consciousness from a neurological standpoint. Last blog post, we defined consciousness via several philosophical standpoints, which was difficult in itself due to the abstract nature of consciousness. While you may think using hard science to define consciousness would therefore be easier, it is still difficult because there is controversy over how to neurologically pinpoint consciousness. Not only is there difficulty in neurologically defining consciousness, but there is also controversy over the best ways to research

consciousness and assess awareness, as we will discuss later. One of the most famous neurological theories of consciousness is Neural Correlates of Consciousness (NCC). This concept connects scientific observations of brain activity to the conscious experiences that participants report. Thus, consciousness is a property dependent on internal, complex neural processes.
For example, EEG is commonly used to study neuroscientific properties of consciousness, which are dependent on the brain’s patterns of neuronal firing and thus its overall electrical activity. When using EEG to study NCC, the main focus lies on the patterns of whole populations, rather than individual, neurons. The goal is to link activity between synchronized neuronal firings in different brain regions (ie - “neural oscillations”) and experiences of consciousness, which was an idea originally proposed by the famous scientists Crick and Kock in 1990. However, one of the first theories about the neurochemistry of consciousness developed a bit earlier in the 1980s. Christof von der Malsburg and Wolf Singer developed the idea that the brain processing and integrating information is linked to gamma oscillations in the brain, which are one of the highest-frequency brain waves. We will discuss in the next section how this principle was later used to connect specific mental states to specific brain wave frequencies.
Other theories for the neural bases of consciousness, besides NCC, are the Global Neuronal Workspace, Information Integration Theory, Recurrent Processing Theory, and Higher Order Theory. The prior two were discussed in the last blog post. As a bit of a recap, Global Neuronal Workspace considers a mental state conscious only when the information it holds is accessible in the “global neuronal workspace” to multiple neuronal systems including perceptual, long-term memory, evaluational, motor, and attentional systems. But how can you tell that this information is actually accessible to these various systems? The answer is simple: accessible information can be used by these systems to carry out their own processing and computations, thus designating that information as conscious information (as opposed to subliminal or preconscious information). Recently, cortical structures in the brain have been proposed as being composed of “workspace neurons” that specifically link neural systems through their long-ranging reach. To recap the second theory, which was also discussed in the previous blog post, Information Integration Theory (IIT) is when one’s neuronal activities and pathways must be able to integrate external sensory information and process it on multiple levels in order to be considered conscious. Finally, the two new neural theories of consciousness I will introduce today are Recurrent Processing Theory and Higher Order Theory. Recurrent Processing Theory

defines consciousness through specific interconnected sensory systems that provide each other feedback for later processing. Recurrent processing is a stage in this series of processing in which information returns from a stage of deeper processing and back to the original sensory area for further local processing. Lastly, Higher Order Theory postulates that someone must be aware of being in a certain mental state for it to be conscious. Thus, one must maintain a higher-order of self-awareness to qualify as conscious, which in this theory is neurologically linked to the prefrontal cortex. The prefrontal cortex, located in the frontal lobe, is linked to the complex logical and personality traits that are associated with being human. However, there are other brain regions linked with higher-level cognitive processing, such as the secondary somatosensory, insular, posterior parietal, and anterior cingulate cortices, which have also become focuses in studies of consciousness using neuroimaging.

While all these theories seem to be wildly different in how they identify the neural bases of consciousness, there is one main controversy that connects them all: whether consciousness originates from the frontal or posterior region of the brain. The posterior region is emphasized in IIT, in which the parietal and occipital areas are a hub for consciousness and integrating
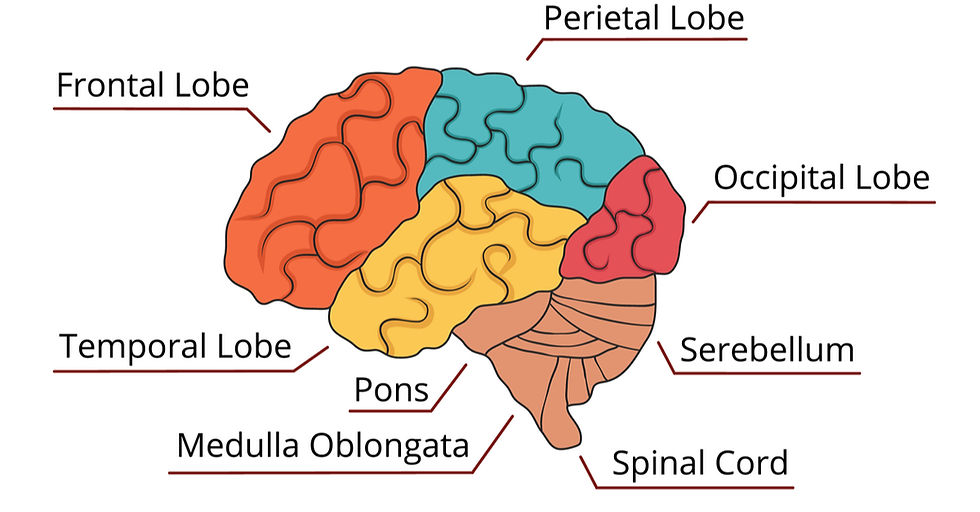
information, and Recurrent Processing Theory, in which the visual pathway is emphasized as a prime example of recurrent processing in the occipital lobe. On the other hand, Higher Order Theory focuses on the frontal lobe. Global Workspace is the only theory that encompasses all of the brain. The prefrontal cortex has been popularly thought of as crucial to consciousness, due to its more advanced functions in humans. One general view within NCC is that prefrontal cortex activation is the true neural correlate of consciousness, while others regard visual activation in the occipital and parietal lobes as the true neural correlate of consciousness.

In recent years, a new brain structure, though, has come to the light as a potential neural correlate of consciousness. This structure is the claustrum: a thin layer of neurons that is ambiguous in its function. In 2017, Christof Koch’s team of neuroscientists first proposed the claustrum, which is located below the cerebral cortex, to be the center that conducts the symphony of neural activity throughout the brain responsible for consciousness. USC researchers led by Jack Van Horn then investigated further into this theory and discovered through neuroimaging that the claustrum is actually very unique in the fact that it has the most physical connections to other brain structures in spite of its small size / volume. This has contributed to speculations about its importance in neural networking across the brain, since sensory input from nearly all brain regions eventually end up at the claustrum. Although it is not a brain structure, working memory is another neural process that has historically been proposed as a neural correlate of consciousness. Working memory, also known as short-term memory, is a cognitive system used to store perceptual and linguistic information for immediate processing.
Clearly, a huge part of consciousness is how we perceive the world. One interesting way to study perception and its relation to consciousness is by studying visual perception, which many people claim to be the focal point of consciousness. There are two visual streams into which primate cortical vision is divided: the dorsal and ventral streams. The dorsal stream, which runs through the parietal lobe, involves spatial perception, particularly of visual motor movements in one’s environment. The ventral stream, on the other hand, which runs through the temporal lobe, involves categorical perception, particularly of conceptual visual stimuli in the

environment. Interestingly, lesions to the dorsal stream do not affect conscious vision. What does that mean? Well, a lesion to the dorsal stream can lead to optic ataxia, a condition that leads to difficulty in motor responses to visual stimuli without the issues of visual perception. In other words, people can perceive visual objects, but are unable to complete tasks like reaching or grasping for the objects in question. Thus, the dorsal stream is often equated to unconscious vision, in that someone unconsciously perceives the object. In contrast, a lesion to the ventral stream does affect conscious vision, and can lead to a condition called visual agnosia. Visual agnosia causes people to not be able to perceive the form of an object or visually categorize / recognize it. Thus, the ventral stream is equated to conscious vision, and is thus tied to normal activity in the primary visual cortex (V1). Another interesting effect of lesions to V1 that give us more clues to NCC is blindsight. People with blindsight maintain the ability to behave and respond to visual stimuli, such as avoiding obstacles, despite the fact that they claim to not consciously see / perceive the stimuli. Recently it has been proposed that blindsight could be attributed to alternative processing pathways such as the lateral geniculate nucleus (LGN) in the thalamus, the pulvinar, and the superior colliculus (SC), which are all relay centers in the visual sensory and processing pathway. The main controversy surrounding blindsight though is whether or not it truly engages unconscious vision. This controversy is fueled by the claim that subjects can minimally detect visual stimuli, but they do not report them as conscious visual perception because they are not confident enough in their limited perception.

To end our discussion on the neural bases of consciousness before we move on to research studies and methods in consciousness, I want to discuss neurostimulation. Neurostimulation, especially in the latter 1900’s, became a popular way to discover the internal neural properties and mechanisms responsible for certain phenomenal experiences. Microstimulation involves delivering a small current directly to nearby neurons via injection, which then can affect other neurons through synaptic connections. We assume that it can affect other neurons because functionally related neurons have synaptic connections, allowing you to influence very specific neural activities and perceptual states in such cases. For example, Wilder Penfield used
electrical microstimulation directly to the primary sensory cortices (V1, S1, etc.) to induce hallucinations in some patients. Alternatively, microstimulation of the Fusiform Face Area, or FFA, a brain region primarily responsible for human face processing, caused visual facial distortions. Similarly, microstimulation of the parahippocampal place area (PPA), which is primarily responsible for spatial navigation, induced visual hallucinations of visual scenes. Not only could microstimulation be used to induce experiences or disrupt normal functioning, but it could also be used to sway perception in specific manners, which is why it is more regulated today. For example, Salzman identified a brain region known as the MT in primates, which, in monkeys, is responsible for discriminating visual motion and movement. By identifying the neurons involved in detecting motion in a particular direction and by microstimulating them, Salzman was able to heighten the monkey’s perception by making them much more sensitive to visual motion in that direction.
Research Methods / The Role of Neuroimaging:
Now that we have discussed how to define consciousness from the standpoint of perception, neural processes, and brain structures, how does one study or even assess consciousness? Neuroimaging has become a very popular tool within the field of consciousness research, since consciousness involves the brain being in a state of arousal that is clearly visible through neuroimagery. However, as has been made clear through our discussion, arousal is not a holistic indicator of consciousness. Under the umbrella of arousal, there are different states of consciousness; for example, sleep EEG patterns are very different than wakefulness EEG patterns, but only the latter state would be considered wholly conscious even though both are states of arousal.


Connecting specific neural networks and pathways to specific information processing pathways and mental states can be difficult. However, researchers develop paradigms, or tests, to help link specific conscious thoughts and information processing to specific brain regions. An example of such a paradigm, which we will discuss in much more depth in the next blog post, was developed by Martin Monti, a consciousness researcher here at UCLA. The paradigm consists of two tasks to detect consciousness in patients in vegetative states; one task asks patients to imagine walking around a room, whereas the other task asks patients to imagine playing tennis. These tasks test the patients’ spatial imagery and motor imagery, respectively, despite them being behaviorally and verbally unresponsive. Dr. Monti thus shows that people can still be perceptive and mentally responsive to their surroundings despite their official diagnosis as “unconscious” or “unresponsive.”

There are lots of neuroimaging tools that are prevalent in neuroscience research, but which are most common in consciousness research? Electroencephalography (EEG), functional magnetic resonance imaging (fMRI), and positron emission tomography (PET) scan are the most common neuroimaging used to assess and study consciousness. Two components of consciousness are wakefulness and awareness, so neuroimaging uncovers how to properly assess and observe these properties in people. EEG tracks electrical activity and neuronal firing patterns in the brain in order to detect neural signatures for different states of consciousness. If you remember one of my earlier blog posts way back in the day, I broke down what mental states are associated with each frequency of electrical activity (alpha = relaxed, beta = conscious / alert, theta = sleep / meditation, delta = deep sleep / characteristic sleep waves, gamma = conscious perception). By observing which frequencies are expressed and/or suppressed on a person’s EEG power spectrum, which graphically shows the contributions of each frequency of brain wave to the overall EEG signal, one can assess what mental state a person is currently in. fMRI, on the other hand, detects changes in blood flow to the brain, and blood flow increases to the brain regions that are active and aroused. PET scans are a bit more versatile in what the image can show you since it uses radioactive tracers that can attach to different compounds. Depending on the compound the radioactive tracer attaches to, a PET scan can show you oxygen use, blood flow to the brain, how the brain uses glucose, and much more. Thus, PET scans can show the mechanisms by which your brain tissue and structures are functioning and whether or not they are functioning properly. Overall, the goal of neuroimaging in consciousness research is to link specific brain patterns and neural activation patterns with certain thoughts, intentions, and states of consciousness in order to assess awareness more scientifically and technically, rather than just relying on behavioral responses. This is especially useful in studying disorders of consciousness, which we will discuss more in-depth in the next blog post. Stay tuned for the next part of this consciousness series, where I will cover a plethora of famous examples of consciousness research design and break down the remarkable discoveries of consciousness research.

Comments